文献介绍

文献题目: 对区域间皮层投射神经元命运多样性的时间控制
研究团队: Denis Jabaudon(瑞士日内瓦大学)
发表时间: 2021-11-09
发表期刊: Nature
影响因子: 49.9(2021年)
DOI: 10.1038/s41586-021-04048-3
摘要
新皮质区域之间的互连对于感觉整合和感觉运动转换至关重要。这些功能由具有异质性的区域间皮质投射神经元 (inter-areal cortical projection neurons, ICPN) 介导,该神经元将轴突分支发送到皮质区域以及皮质下目标。尽管 ICPN 在解剖学上具有多样性,但它们在分子上是同质的,并且它们的解剖学和功能特征的多样性在发育过程中如何出现仍然很大程度上未知。在这里,作者通过使用 MAPseq 和 scRNAseq 关联发育中小鼠新皮质的单个 ICPN 的连接组 (connectome) 和转录组 (transcriptome) 来解决这个问题。关注于初级体感皮层 (S1) 的神经元,作者揭示了运动皮层投射( ) ICPN 与次级体感皮层投射( ) ICPN 的分子和功能分化的长期展开。作者将 SOX11 确定为 相比于 暂时的差异表达转录因子。出生后对 S1 中 SOX11 表达的操纵会损害小鼠的感觉运动连接并扰乱选择性探索行为。总之,作者的结果表明,在单个皮质区域内,ICPN 的不同亚型具有不同的出生后分子分化速度,随后反映在不同的环路连接和功能中。因此,一组基本通用的基因表达水平的动态差异,而不是发育遗传程序特性的根本差异,可能解释了皮层神经元内部类型多样性的出现。
研究结果
来自外周的体感输入通过丘脑到达初级体感皮层 (S1),信息从这里传递到其他皮层区域和皮层下目标进行进一步处理。在皮层内,S1 与运动区 (M) 和次级体感区 (S2) 紧密相连,形成感觉运动协调(S1-M 连接)和感觉辨别(S1-S2 连接)的平行通路。在成人中,M- 和 S2- 投射 ICPN(此处分别称为 和 ICPN)对这两个区域的投射在很大程度上是相互排斥的,但这种特殊性在发育过程中如何出现尚不清楚。作为对这个问题的第一次尝试,作者检查了 S1 中 和 ICPN,在出生后 P5(当丘脑皮质突触在 S1 中形成时)、P7 和 P14(当 S1 区域间投射基本建立时)使用逆行标记(Fig. 1a, Extended Data Fig. 1a, b)。这种方法表明,从 P5 开始,对 M 和 S2 的投射在很大程度上是相互排斥的。在那个年龄时期,只有极少数逆行标记的 和 ICPN 在浅层 (superficial layers, SL) 中可见(即,大多数轴突尚未到达其目标;Fig. 1a, Extended Data Fig. 1a, b),可能反映了先是深层 (DL) 然后是浅层神经元的顺序诞生和发育。因此,在出生后第一周,皮质感觉运动功能可能最初由深层 ICPN 驱动,而浅层 ICPN 在后期发育阶段发挥作用。
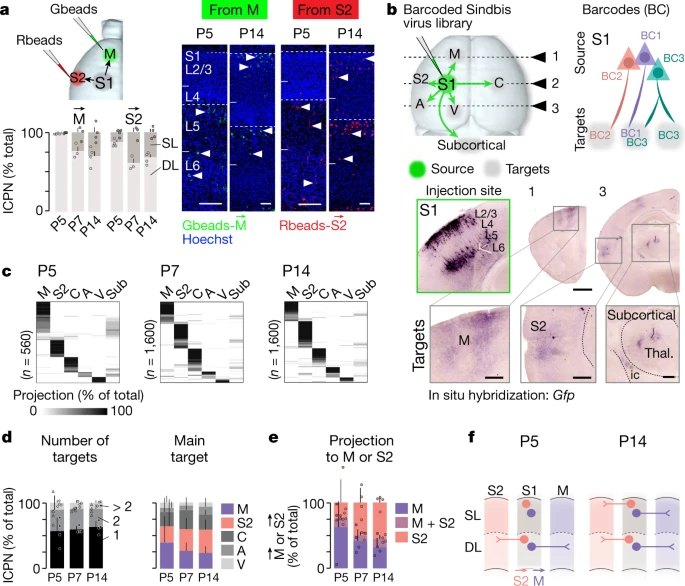
a. 左上图,使用绿色(Gbeads)或红色(Rbeads)retrobeads 从 M 或 S2 逆行标记。右图,S1 中 P5 和 P14 时期 Gbeads 标记的 M→ ICPN 和 Rbeads 标记的 S2→ ICPN(箭头表示逆行标记的 ICPN)。左下图,P5、P7、P14 时期浅层(SL)与深层(DL)相比 M→ ICPN 和 S2→ ICPN 的分布 (P5: n = 4, P7: n = 3, P14: n = 4 pups from two litters per target)。
b. 上图,MAPseq 原理。下图,展示了 P14 时期投射到靶点的轴突 barcode-Gfp mRNA 原位杂交。绿框图像:注射位点。
c. 单个 S1 ICPN 在 P5、P7、P14 时期的轴突投射,数字表示 ICPN 的数目。
d. P5、P7、P14 时期 ICPN 的投射个数(左)和主要投射区域的比例(右)。
e. P5、P7、P14 时期 ICPN 投射到 M、S2、M + S2 的比例。
f. 在出生后的发育过程中,S1 投射到 M 和 S2 的示意图。
然后,作者使用 MAPseq 来绘制 S1 中单个 ICPN 中多个轴突投射的 target 特异性发育。MAPseq 使用来自胞体的 barcoded RNA 的顺行运输,实现对多个远程 targets 的轴突投射的高通量单细胞重建(Extended Data Fig. 1c, Supplementary Note 1)。检查了六个功能相关 targets:ipsilateral M、S2、auditory (A)、visual cortices (V)、contralateral S1 (C)、subcortical targets (Sub)(即,纹状体和丘脑)在 P5、P7 和 P14 时期(Fig. 1b, Extended Data Fig. 1d)。MAPseq 映射显示,在所有年龄段,M、S2、C 是主要的皮质内目标。 和 ICPN 约占所有年龄段 S1 ICPN 的 70%,其中约 30% 的神经元具有多个 targets;然而 和 ICPN,共享基本相同的 targets 组合(Fig. 1c, d, Extended Data Figs. 1e–h, 2, Supplementary Table 1)。与逆行标记的情况一样,同时投射到 M 和 S2 的一小部分神经元随着时间的推移保持稳定,证实了 和 ICPN 很早就被指定成这样(Fig. 1e, Extended Data Fig. 1b)。总之,这些数据表明,一旦可检测到投射,S1 ICPN 就与 M 和 S2 具有稀疏且定向的连接(Fig. 1f)。
为了研究 和 ICPN 的发育转录动态,作者将 MAPseq 映射与 S1 神经元的 scRNA-seq 结合起来,从而将 barcode-identified 神经元的转录身份与其相应的投射联系起来,并将这种方法称为 ConnectID(Fig. 2a, Extended Data Figs. 3, 4, Supplementary Note 2)。作者在 P7、P9、P14 进行了 ConnectID(在 P5 时期,只有少数浅层 ICPN 轴突到达 M 和 S2;Fig. 1a),使作者能够识别 ICPN 的不同亚型,包括 和 ICPN,及其相应的转录特性(Fig. 2b)。虽然 ICPN 分子身份与神经元年龄明显相关(Fig. 2b,左),但它似乎与连接性无关(Fig. 2b,右)。因此,正如成年期的情况一样,尽管它们有不同的连接性, 和 ICPN 在出生后发育过程中不能被识别为不同的分子细胞类型(Extended Data Fig. 5)。
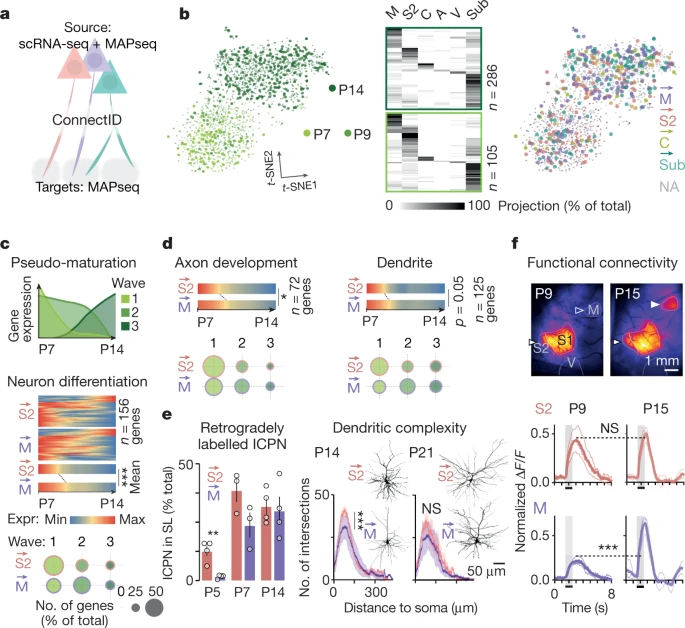
a. ConnectID 将单个神经元中的转录组与投射组连接起来。
b. 左图,t-SNE 表示 scRNA-seq ConnectID(P7, P9 and P14; n = 2,134 neurons)和逆行标记(Rbeads) M→ 和 S2→ ICPN (P9; n = 133 M→, n = 183 S→ ICPN)数据集。大点,targets 中带有 barcode 的神经元(n = 391 ConnectID neurons)。中间图,ConnectID 神经元的多重轴突投射。右图,ConnectID 神经元(大点)和 Rbeads M→ 和 S→ ICPN(小点)的主要 target。灰点,未指定的 (NA) 神经元(即,在任何 target 中均未检索到 barcode 的神经元)。
c. 上图,基因表达动力学(kinetics)揭示了三种转录波(waves)。下图,S2→ 和 M→ ICPN 中每个波中单个基因表达动力学、与神经元分化相关的基因数量。与 S2→ ICPN 相比,M→ ICPN 中与神经元分化相关的基因在表达时间延长。
考虑到 和 ICPN 之间的分子重叠 ,作者假设 ICPN 多样性可能源于以不同速度展开的大部分通用转录程序,而不是源于这些程序性质的根本差异。为了检测 和 ICPN 相应基因集表达水平的潜在时间差异,作者使用单细胞 pseudotime alignment 来绘制它们的发育表达(Extended Data Fig. 6a)。该方法确定了 和 ICPN 中三个连续的基因表达波:第一个波是早发型(P7),第二个波中的基因在整个出生后年龄都有持续表达,第三个波是晚发型(P14)(Fig. 2c, top, Extended Data Fig. 6b)。功能与神经元分化相关的基因显示出在 ICPN 中的延长表达,相比于 ICPN(Fig. 2c, bottom, Extended Data Fig. 6c, Supplementary Table 2)。这种转录转变在第三个波的基因中不再可见,表明在出生后的前两周内发生了一个短暂的过程。时间转移的基因集包括具有与轴突发育和树突相关功能的基因(Fig. 2d, Extended Data Fig. 6c-f, Supplementary Table 3),表明潜在的功能相关性。支持这种可能性, ICPN 表现出轴突延伸和树突树成熟延迟,相比于 ICPN(Fig. 2e)。因此,与 ICPN 相比, ICPN 具有延迟转录程序,导致形态分化相对较长。
接下来作者检查了 和 ICPN 基因表达和解剖特征的时间差异是否存会影响 S1-to-M 和 S1-to-S2 连接的功能成熟度。为此,作者使用了 Cux2-cre × GCaMP6s 转基因幼崽(Cux2 在 SL ICPN 中表达),并对胡须垫、前肢和后肢刺激时的皮质激活进行了体内宽视野成像(Fig. 2f, Extended Data Fig. 7a)。作者设计了成像设置,以便可以在同一视野中监测 S2、S1 和 M 区域,并在 P9(当幼崽开始探索其环境时)和 P15(当探索已充分建立时)进行记录。作者发现,在 P9 时,S2 激活已经达到了 P15 时的响应强度,但 M 激活的情况并非如此,M 激活要弱得多。这揭示了出生后前两周内先是 S1-to-S2 通路随后是 S1-to-M 通路的连续功能发育(Fig. 2f, Extended Data Fig. 7b, c)。总之,这些数据共同表明 M→ ICPN 具有分子、解剖学和功能发育延迟,相比于 S2→ ICPN。因此,即使在单个区域和层内,密切相关的神经元亚型也表现出不同的转录和相关的出生后功能成熟速度。
接下来作者测试了对时间调节转录程序的操作是否会影响 S1 的区域间连接。为此,作者计算了 和 ICPN 之间所有 wave 1 genes(即,轴突发育时高度表达的基因)沿着 pseudo-maturation 的基因表达距离(Fig. 2d, Supplementary Table 4)。着眼于转录因子和轴突发育相关的转录本,作者将 Sox11 确定为一个有趣的候选因子,因为该转录因子控制视网膜神经节细胞中的轴突引导,并且在出生后的浅层(SL)神经元中特异性表达(Fig. 3a,b)。Sox11 转录和蛋白的表达水平在 P7 和 P14 之间随着时间的推移而降低,但在 ICPN,Sox11 最初高表达并迅速下降,而在 ICPN,初始表达较弱(Fig. 3b);M 和 S2 逆行标记后使用免疫组织化学证实了这一点(Extended Data Fig. 8a)。为了研究 SOX11 在出生后 ICPN 发育中的潜在作用,作者通过在胚胎期 E15.5 子宫内电穿孔结合 Cre 依赖性质粒,在 S1 的 L2/3 中特异性过表达 Sox11 基因(以 L2/3 ICPN 为目标)在 P0 的 S1 中立体定向注射腺相关病毒 AAV-pCMV-Cre(Fig. 3c, Extended Data Fig. 8b, c)。结合 M 和 S2 的逆行标记,这种方法揭示了 SOX11 过表达 ICPN 中几乎完全不存在 S1-to-M 的投射(Fig. 3d)。 ICPN 标记的减少并未反映细胞死亡或错误迁移,正如与对照相比,P17 时表达 SOX11 的 L2/3 神经元数量相似所证明的那样(Extended Data Fig. 8d)。相反,这个结果可以反映(1) ICPN 轴突重新路由延伸至其他 targets 或 (2) 这些神经元的轴突延伸减少。对于(1),顺行轴突信号的量化并未显示对 S2 的投射增加(事实上 S1-to-S2 的投射也减少了,尽管不太明显;Fig. 3e, Extended Data Fig. 8e)。同样,对侧皮层的投射或皮层下的投射也没有增加(Extended Data Fig. 8e)。对于(2),白质内的投射没有受到可检测到的影响(Extended Data Fig. 8e),这表明轴突能够投射到 S1 之外,但随后不会侵入它们的皮质目标。因此,体外 SOX11 过表达 ICPN 中轴突生长并未受到干扰(Extended Data Fig. 8f),表明 SOX11 本身不会损害轴突延伸,而是可能调节对靶标衍生分子线索(cues)的反应。与 SOX11 在轴突引导中的重要作用一致,使用可诱导的 CRISPR–Cas9 Sox11 单引导 RNA (sgRNA) 质粒构建体从 S1 ICPN 中的 P0 进行条件消融导致 S1 的皮质内投射总体减少(Extended Data Fig. 8g) 。 ICPN 投射受这种操作的影响较小,这与这些神经元中 SOX11 表达的基线水平较低一致。因此,SOX11 对于 ICPN 轴突侵入其皮质目标是必需的。总体而言,这些数据表明出生后 SOX11 表达水平对于正确的区域间感觉运动环路组装至关重要。
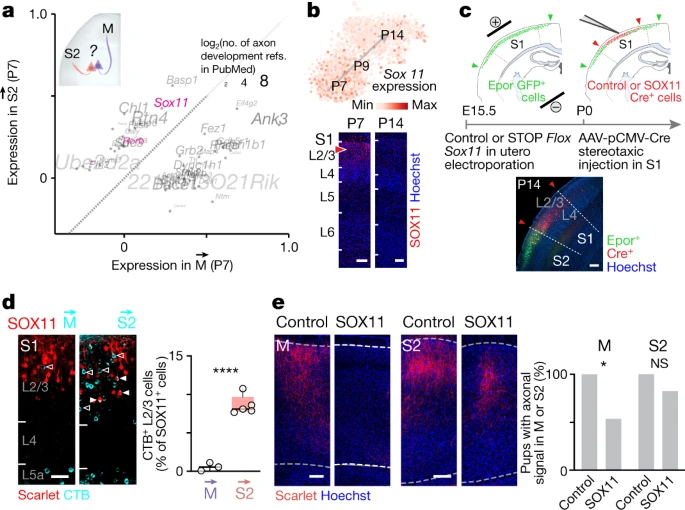
a. P7 时 和 ICPN 之间的差异基因表达,此时轴突到达 target。P7、P9、P14 ConnectID 神经元的总数(用于计算 和 ICPN 之间所有 wave 1 genes 沿着 pseudo-maturation 的基因表达距离)n = 114 ; n = 76 ICPN。每个基因符号的大小代表带有术语“轴突发育”的 PubMed 摘要数量。这里只展示了具有超过一篇与轴突发育相关的 PubMed 摘要的基因。转录因子以粉红色显示。
b. Sox11 在出生后的发育过程中动态表达(上图),在蛋白水平上得到了证实(下图)。
c. 在 E15.5 的子宫电穿孔结合 AAV-pCMV-Cre 立体定向注射推测 P0 时 S1 的 Sox11 空间和时间限制的过表达。
d. P15 时期 M 或 S2 的逆行标记,以及 P17 时期 SOX11-overexpressing (SOX11) 的 S1 L2/3 M→ 和 S2→ ICPN 胞体的定量 (M: n = 3; S2: n = 5 pups from 2 litters)。
e. S1 Cre+ 轴突在 P17 的 M 和 S2 中表达猩红(Scarlet)。左图,每个 target 中 Scarlet+ 轴突的代表性图像。右图,基于荧光的轴突投射定量。
接下来,作者检查了 S1 中 SOX11 单侧过度表达后感觉运动连接中断的行为后果(Fig. 4, Extended Data Fig. 9)。当在开放视野场地进行测试时,S1 SOX11 过表达小鼠的空间探索受到严重损害(Fig. 4a, b, Extended Data Fig. 9a, b)。这可能反映了真正的感觉运动障碍,而不是与焦虑相关的行为,因为在开放区域中心花费的时间与对照小鼠相似(Extended Data Fig. 9b)。SOX11 过表达小鼠的触觉空间探索是强烈不对称的,它们大部分时间都花在探索目标 S1 对侧的墙壁上(Fig. 4c)。总体而言,行为改变与 S1 连接中断的程度相关(Extended data Fig. 9c, d),表明与感觉运动探索存在直接因果关系。
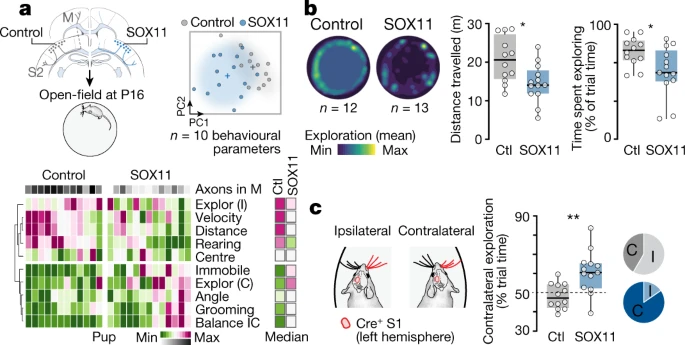
a. 左上,对照 (ctl) 和 SOX11 过表达 (SOX11) P16 幼崽在开放视野场地中监测 10 分钟。主要探索参数的主成分分析(右上)和热图(下)。C,对侧;Explor,探索;I,同侧。
b. 左图,所有 control 和 SOX11 过表达的受监测幼崽的平均轨迹。右图,量化行进距离(unpaired t-test, P = 0.0137)和探索时间(unpaired t-test, P = 0.0227; n = 12 control pups, n = 13 SOX11 pups)。
c. 花时间探索电穿孔 S1 的同侧和对侧开放场壁。饼图代表花费时间靠近同侧或对侧墙壁的幼崽的比例(n = 12 control pups, n = 13 SOX11 pups; paired t-test, control: P = 0.360; SOX11: P = 0.0073; 大多数时间幼崽在同侧与对侧壁上的消耗:FFisher’s exact test, P = 0.0414)。
作者的结果表明,转录相似的神经元以不同的方式连接的策略是让它们以不同的速度成熟。具体来说,对共享基因表达水平的动态控制可能提供一种简约的机制,以在发育过程中生成跨转录同质的 ICPN 亚型的一系列投射。由不同时间调节因子调整的“通用”转录程序的重新利用可能是因其代谢适应性而被选择的,从而能够实现探索行为所需的区域间网络的快速和稳健的连接。细粒度的时间剂量依赖性控制可能允许与不同的细胞外分子梯度相互作用,最终影响轴突引导。支持这一过程的是,在真兽类小鼠与有袋动物 dunnart 中,主要共享转录程序的时间差异是胼胝体与半球间轴突前连合交叉的基础。除了这些时间控制之外,在发育的早期阶段(包括在胚胎中),独特的和确定性的分子程序可能在 和 ICPN 启动轴突目标特异性并诱导此处确定的时间移位。
就行为而言,积极的环境探索在生命的前两周逐渐出现,这对应于此处确定的 S1-to-M 连接的长期成熟。因此,交错发展 ICPN 和 ICPN 可以为感官技能(例如,母乳喂养最初所需的技能)和运动技能(例如,漫游所需的技能)的交错发展奠定基础,并帮助在感觉输入和运动学习所需的协调运动输出之间建立顺序关联。
注:本文为个人学习笔记,仅供大家参考学习,不得用于任何商业目的。如有侵权,请联系作者删除。

本文由 mdnice 多平台发布




![[数据集][目标检测]街头摊贩识别检测数据集VOC+YOLO格式758张1类别](https://i-blog.csdnimg.cn/direct/661724e63a6b435d8806320704b39ea8.png)